
Motor Control
last authored:
last reviewed:
Introduction
One of the principle functions of generating internal representations of the world is to guide movement. Even simple actions require sensory information about what and where our actions are. Proprioreceptive information is needed.
Brain Control
Planning and Initiation
Motor plans are thought up by the posterior parietal and premotor areas. The premotor cortex specifies spatial characteristics of movement based on sensory information from the parietal cortex.
The purpose of movement is determined by the dorsolateral frontal cortex. Following stroke in some areas, volitional control is gone and someone can't smile symmetrically if told, though a joke will cause a symmetrical smile.
Homunculus, courtesy of btarski
Primary motor cortex
- much devoted to the face and mouth (speech) and the hand
- very little for the rest of the body
pre-motor cortex
supplementary motor cortex
- involved in complex movements, both contralaterally and ipsilaterally; also engaged during mental rehearsal of complex movements
Processing
Many structures are involved with processing motor control.
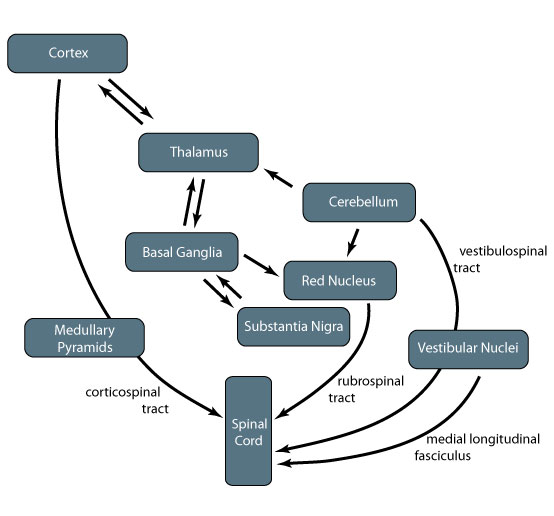
Basal ganglia mediates many of the feedback loops involving other components of motor control. Parkinson's disease is caused by neurodegeneration of cells in the basal ganglia, resulting in motor symptoms such as weakness, poor initiation of movement, and tremor.
The thalamus is a central junction as information passes from the cerebellum, basal ganglia, subthalamic nuclei, and substantia nigra to the motor cortex. It also sends information back to the striatum in the basal ganglia.
Central Pattern Generators
Central pattern generators (CPGs) are neural circuits set in place before movement begins that can be sent to muscles with appropriate timing. This removes the need for sensory feedback during a sequence of movements, facilitating activity. They mediate behaviours that include walking, running, breathing, chewing, certain eye movements, and even scratching. Central pattern generators for breathing, initiation, and speed reside in the brain stem, while those for locomotion are present in the spinal cord.
The fact that a chicken with its head cut off will run around shows that central pattern generators are present in the spinal cord.
At the core of CPGs is a set of cyclic, coordinated timing signals generated by a cluster of interconnected neurons. These affect as many as several hundred muscles that precisely contract or relax during a movement.
To achieve coordination among the limbs, sets of CPGs are interconnected. They are also very flexible, turning on or off as needed or adapting to variables such as terrain.
There is no single way to generate rhythmic patterns, and different mechanisms are used. The simplest are single neurons with membrane properties making them pacemaker cells, similar to cardiac nodal cells. These cells are embedded within interconnected circuits.
Other, more complex models of pattern generation include the half-centre model, which suggests two excitatory interneurons are connected to reciprocally inhibit one another. This leads to rhythmic cycling from one center to another.
Sensory feedback, in the form of stretch receptors, acts to terminate activity on one side and initiate it on the other. Spinal networks also reciprocally communicate with the brainstem to maintain control capacity in higher centres.
Motor Neurons
Motor neurons are massive cells, with a huge dendritic tree, located in the brainstem or the ventral horn of the spinal cord. Each lower motor neuron innervates muscle fibres of the same type.
- α motor neurons innervate the main force-generating muscle fibres (extrafusal fibres)
- γ motor neurons innervate only fibres of muscle spindles.
Upper motor neuron is a term used clinically to indicate a population of neurons with oligosynaptic input to lower motor neurons. These usually refers to cells in the brain, but can also include those of the spinal cord.
Lower motor neurons originate in the brainstem or spinal cord and leave the spinal cord to directly synapse to innervate muscles.
Distinguishing Upper vs Lower Motor Neuron Problems:
UMN |
LMN |
|
muscle bulk |
normal |
atrophy |
muscle tone |
increased (spasticity) |
normal or decreased |
tendon reflexes |
increased |
normal or decreased |
Babinski |
positive |
normal |
other |
- |
fasciculations |
Spinal Pathways and Output
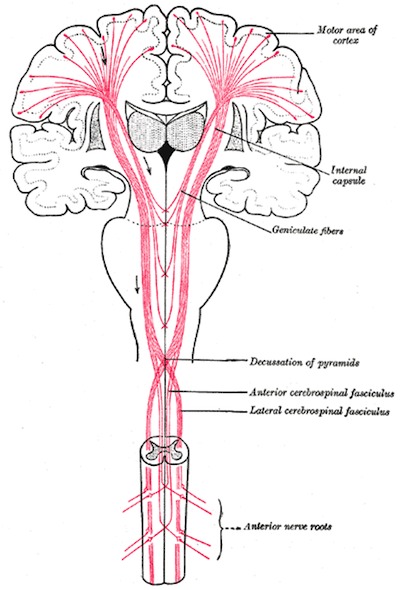
Upper motor neurons, from Gray's Anatomy
Descending pathways synapse in the medial and lateral alpha motoneuronal nuclei. Medial nuclei are found at almost every level, while lateral nuclei are prominent in the upper and lower limb cord enlargements. There is some predictable anatomical correlation (somatotopy) here, with flexors being more anterior and extensors being posterior.
Descending motor tracts can synapse in the spinal cord, mediating activity there, or exit the central nervous system through the ventral horn to innervate muscle.
The spatiotemporal details of muscle contractions required to execute planned actions are coordinated by motor circuits in the spinal cord.
Corticospinal tract
Motor commands for fine, learned, complex movements (ie finger/hand) arise in the motor cortex (30%); pre-motor cortex (30%), and parietal (somatosensory) cortex (40%), and descend through the corticospinal tracts.
Axons descend ipsilaterally through white matter of the internal capsule and the cerebral peduncles of the midbrain, breaking into fascicles as they enter the basilar pons. They then descend through pyramidal tracts of medulla, where 85% of fibres cross over to form the lateral corticospinal tract (85%), which extend the length of the spinal cord. The other 15% do not cross and descend in the anterior corticospinal tract. These upper motor neurons synapse in the ventral horn upon αlpha motor neurons, which exit along the ventral root.
Corticobulbar tract
The corticobulbar tract carries motor information from the cortex alongside the corticospinal tract, terminating on the reticular formation and on motor nuclei of the cranial nerves.
Medial longitudinal fasciculus
The MLF is responsible for coordinating head and eye movements. It begins in the vestibular nuclei and runs along the midline of the brainstem to the cervical spinal cord, where it synapses on interneurons. Some fibres end in the abducens, trochlear, and oculomotor nuclei.
Problems in the MLF can lead to internuclear ophthalmoplegia, where horizontal gaze is disrupted in one eye past midline.
Rubrospinal tract
The rubrospinal tract arises in red nucleus in midbrain, which receives input from deep cerebellar nuclei and motor cortex.
Reticulospinal tract
The reticulospinal tract has projections from the cortex to the reticular formation; these descend in lateral column to terminate on spinal cord interneurons and gamma motor neurons.
Vestibulospinal tract
The vestibulospinal tract arises in vestibular nuclei, which receive input from the vestibular nerve and cerebellum. The lateral fibres innervate alpha and gamma motor neurons all along spinal cord, playing an important role in maintaining posture. The sensation that follows rapid spinning, with symptoms of vertigo and gait instability, is the product of overactive vestibulospinal tract. The medial tract innervates the cervical spinal cord and is responsible for head position. It runs alongside the medial longitudinal fasciculus (described above).
Tectospinal tract
The tectospinal tract arises from cells in the superior colliculus of the midbrain to controls reflex movements of upper trunk, neck, and eyes in response to visual stimuli.
Some descending fibres become incorporated into the medial longitudinal fasciculus in the medulla, while other fibres descend in anterior funiculus of spinal cord and terminate at the cervical level, where they synapse on motor neurons.
monoamines modulate how motor neurons fire, in order to change the signals which are required to maintain muscle strength.
Motor Units
All the neurons that innervate a single muscle form a motor neuron pool.
Each motor neuron commands a group of skeletal muscle cells. Each muscle cell belongs to only one motor unit, which can vary in size depending on muscle function. Extraocular muscles can have just a few fibres per unit, while some leg muscles can have several thousand muscle fibres per motor unit.
Resources and References
 |